

Pigs are the only domestic species naturally infected by the ASFV. European wild boars are also susceptible to infection, showing similar clinical signs and mortality to domestic pigs. However, African suidae usually have subclinical and non-apparent infections, thus acting as reservoirs of the disease.
African Swine Fever virus also replicates in Ornithodorus moubata ticks in Africa and Ornithodorus erraticus ticks in the Iberian Peninsula, both of which are biological vectors and virus reservoirs.
ASFV is the only DNA virus amongst the arboviruses.

O.erraticus, present in the Iberian Peninsula

O. moubata participate in the epidemiologic cycle of ASF
in the South and East of Africa.
Picture by The Natural History Museum of London, 2005.
There are significant epidemiological differences between Africa and Europe. In Africa, ASFV is maintained by a cycle of infection between African wild boars and the biological vector O. moubata, present in several regions in sub-Saharan Africa, so it is difficult to eradicate. Clinical signs and mortality are observed only when domestic pigs are present.

Warthog (Phacochoerus aethiopicus). This is one of the most frequent
wild African suinae species. Picture by Carmina Gallardo
The virus multiplies in the intestine of Ornithodorus and emigrates to the saliva glands and reproductive organs, spreading from males to females during copulation. In the case of O. moubata, transovarian transmission to the progeny also occurs.
In contrast, European wild boars are susceptible to ASFV infection. But in this case, the clinical signs and mortality are similar to those observed in domestic pigs. The epidemiological dynamics of boars and domesticated pigs is also very similar, the most common infection route being direct contact between sick and healthy animals. Indirect transmission via biological vectors such as O. erraticus has been described in the Iberian Peninsula, especially in extensive farming regimes.
Other significant differences between Africa and Europe are related to ASFV and biological vectors. In Africa, O. moubata transmits the virus via the transovarian and transestadial routes whereas in Europe transestadial transmission has only been observed in O. erraticus.
Traditionally, it enters virus-free regions via uncooked pork waste fed to pigs, especially from ships and aircraft.
Once the disease is established in a country, it mainly spreads through direct contact between sick and healthy animals. Recovered carrier pigs play a fundamental role in transmitting the disease.
The virus can be indirectly transmitted via lorries, drinking and eating troughs, surgical and personal equipment, rodents, other animals on the farm, etc, as well as via biological vectors.


Electron microscope image where the ASFV associated
with erythrocyte can be observed.
Picture by J. Quintero
| Main factors involved in ASF transmission |
| Direct contact between healthy and sick pigs. |
| Carriers |
| Indirect route: Transport, drinking and eating troughs, surgical equipment, personal equipment (clothes, footwear), rodents, other animals on the farm, etc, and also biological vectors. |
| Main routes of ASFV elimination |
| Nasal secretions |
| Saliva |
| Faeces |
| Urine |
| Conjuntival exudate |
| Genital exudate |
| Bleeding wounds |
Virus elimination starts even before the first signs appear, during the incubation period. In the acute stage of infection the virus titre in tissues and blood is very high and viraemia is maintained during convalescence.
ASFV persists for several weeks or months in uncooked or frozen meat.
In cured meat such as Parma ham, the virus is inactivated from 300 days after curing has finished. In Spanish products such as cured ham, cooked ham and shoulder, and pork loin, the ASF survival period is shorter than that established for the commercial curing of each product.
| Inactivation of the ASF virus in pork products | ||
| Curing time (days) | ASF virus inactivation time (days) | |
| "Serrano" cured | 180-365 | 140 |
| Iberian ham | 365-730 | 140 |
| Iberian pork shoulder | 240-420 | 140 |
| Iberian pork loin | 90-130 | 112 |
AFRICAN SWINE FEVER PATHOGENY:
1. ASFV usually enters via the oronasal route, though the cutaneous (as a result of scarification), intramuscular, subcutaneous and intravenous routes (tick bites) are also possible. The incubation period varies between 3 and 21 days, depending on the isolate and exposure route.
2. Viruses spread from the monocytes and macrophages in the lymphatic ganglia via the blood, associated to the haematic membranes and/or via the lymphatic route.
3. Viraemia usually begins 2-3 to 8 days post-infection (dpi) and, due to the absence of neutralising antibodies, persists for a long time, even months. Second replication occurs as the the ASF virus reaches the different organs, for example the lymphatic ganglia, medulla ossium, spleen, kidneys, lungs and liver.
No infectious virus is found in ham boiled or cooked at 70 °C.
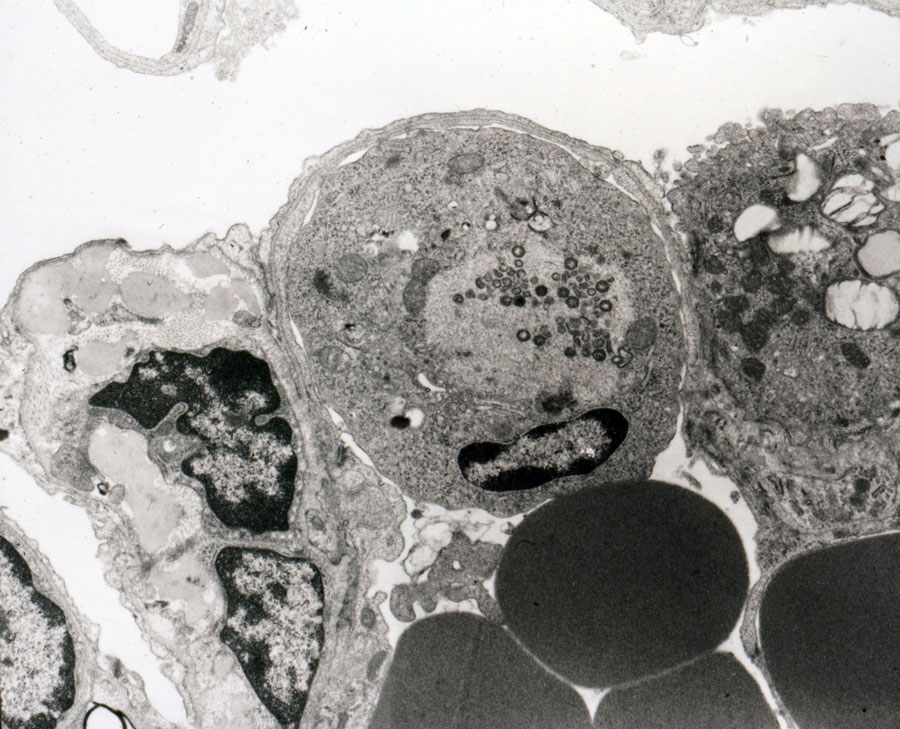
Macrophages infected by ASFV
The main target cells of the ASF virus are those belonging to the Mononuclear Phagocytic System (MPS) which undergo a cytopathic effect characterised by a rounding of the nucleus, peripheric margination of the chromatin and vacuolisation of the vacuolar system culminating in necrosis of the cell with viral replication. One of the consequences of this cytopathic effect is haemadsorption (adsorption of the porcine red corpuscles surrounding the macrophages infected by the virus), which is considered responsible for passive transport of the virus by the erythrocytes.
The ASF virus not only induces necrosis of MPS cells but also intense activation and release of chemical mediators of inflammation by these cells, mainly in the initial stages of the disease. Although the monocyte-macrophage is the main ASFV target cell, replication has been described in other cell populations, so it is currently considered to be a pantotropic virus.
The red splenic pulp in pigs is formed by venous sinuses and Billroth's cords. The Billroth's cords form a tridimensional network with trabecules of smooth muscular fibres coated in macrophages, which is why the spleen of pigs has a fixed macrophagous population.
During the first days of infection with highly virulent isolates intense viral replication occurs in these macrophages which, like other macrophagous populations, undergo the cytopathic effect produced. The basal membrane remains uncovered and activated platelets and fibrin networks bind to it, causing coagulation throughout the entire spleen. Coagulation immediately stops the return blood circulation while the arterial blood continues to enter, therefore causing hemorrhagic infarction. Haemorrhagic infarction may be incomplete, or even absent in infections caused by moderately virulent strains because the cytopathic effect of the virus is lower.
Haemorrhaging and splenic lesions are the most characteristic and constant lesions in ASF. Several hypotheses have been put forward to explain the pathogenic mechanism that brings about a final state of "shock", combining the direct action of the virus on the endothelium and the disseminated intravascular coagulation (DIC), but the establishment of these processes occurs prior to ASF virus replication in the endothelial cells and this was rejected as the origin of haemorrhaging. However, the appearance of haemorrhaging coincides with intense fagocytic activation of the endothelial cells, increased fenestrations in the capillary walls, the existence of subendothelial deposits and loss of endothelial cells, which leads to the basal membrane being exposed, the platelets being activated and to an intravascular coagulation process being triggered.
These endothelial lesions are the morphological manifestation of bio-chemical changes indicating a dysfunction such as high levels of factor VIII and decreased prostaglandin values. The cytokins released by the activated monocytes and macrophages may also play an important role in establishing DIC, as these cells have been observed in the vicinity of the damaged blood vessels. However, in subacute courses in which haemorrhaging is more intense and widespread, there is no endothelial damage or disseminated intravascular coagulation. Haemorrhaging in these courses is associated with intense vasodilatation and increased vascular permeability, which also justifies the appearance of interstitial oedema.
The increased number of lymphocytes in apoptosis in acute courses of ASF has recently been considered responsible for lymphopenia observed in these forms of the disease. This programmed cell death would be responsible for lymphopenia, initially made worse by the inhibition of the lymphocyte proliferation induced by the virus, and, in the final stages of the disease, by the destruction of lymphoid structures in the spleen, lymphatic ganglia and tonsils as a result of vascular damage. Several mechanisms may trigger apoptisis in viral diseases; in the case of ASF this is attributed to the synthesis and release of different monokins by activated monocytes-macrophages and/or macrophages with viral replication.
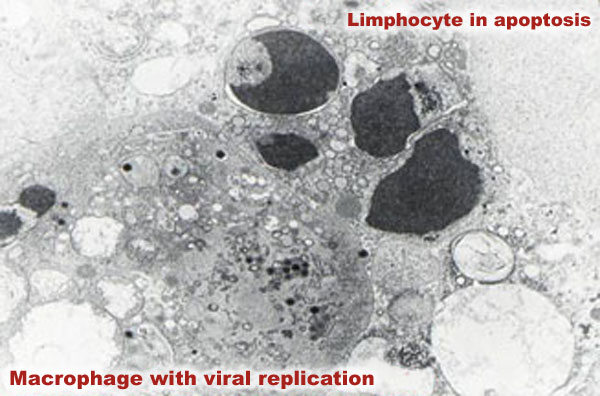
The fact that apoptosis is more widespread in the diffused lymphoid tissue, where there are more activated macrophages, with or without sites of viral replication, supports this hypothesis; paradoxically, the ASF virus contains a gene that inhibits apoptosis in the cells where it replicates.
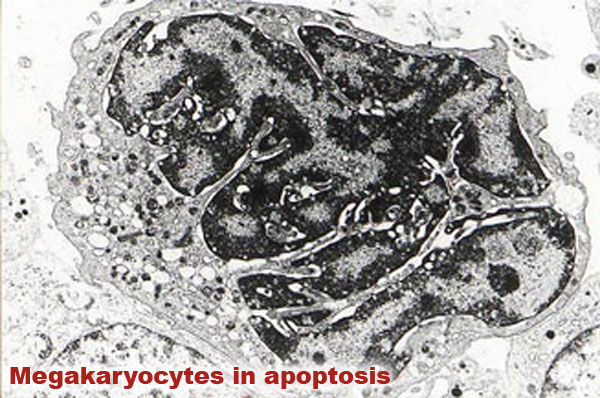
In acute forms of ASF, thrombocytopenia appears in the final stage of the disease, when haemorrhaging is very evident, though it often goes unnoticed because the animals die, which is the result of serious damage in the medulla ossium due to viral replication in the stromal and haematopoietic cells. However, transitory thrombocytopenia in the initial and middle stages of the subacute form of the disease is characteristic. It actively participates in the appearance of haemorrhaging and can cause death if the animal does not overcome this stage of thromboyctopenia.
In subacute courses the appearance of thrombocytopenia is related to three stages or phases of megakaryocytes: a "compensating" stage characterised by the presence of numerous cytoplasmatic projections in the megakaryocytes, which coincides with peripheric consumption of platelets in the initial stage of the disease; a "hypermature or exhaustion" stage with abundant megakaryocytes in apoptosis (also called "naked megakaryocytes"), which is not related to viral replication in these cells and which would cause sudden and serious thrombocytopenia as platelets are not released into the blood circulation; the third stage is a "regenerative" stage where nests of cells are observed in which replication occurs. This indicates a recovery of the population of megakaryocytes and that thrombocytopenia no longer exists.