
Возбудитель АЧС – крупный оболочечный ДНК-содержащий вирус с икосаэдральной симметрией капсида. Его морфология и свойства очень сходны с таковыми у Иридовирусов, а организация генома сходна с Поксвирусами. Недавно вируса был выделен в отдельной семейство Asfarviridae, род Asfivirus, и является его единственным представителем.
Вирусные частицы имеют диаметр 200 нм и состоят из трех концентрических оболочечных слоев, покрытых внешней шестиугольной оболочкой, которая формируется в результате почкования через мембрану инфицированной клетки.
Шестиугольный икосаэдрический капсид окружает ядро диаметром 80 нм, которое, в свою очередь, окружено липидной оболочкой.
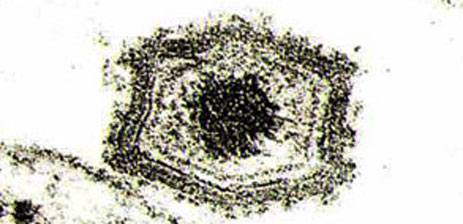
Вирионы АЧС электронная микроскопия
| Характеристики вирусной частицы |
| Средний размер частицы - около 200 нм |
| Электронноплотное ядро жиаметром 80 нм |
| Как минимум три оболочки, и еще одна у внеклеточного вируса |
| Характеристики вирусного генома |
| Молярная масса: 100,1 г/моль-1 |
| Плотность в ClCs:: 1,7 г/см3 |
| Коэффициент седиментации: 60s |
| Длина: 58 нм |
Вирусные частицы неустойчивы к тепловому воздействию и чувствительны к жировым растворителям, но очень устойчивы в широком диапазоне pH.
Репликация вируса происходит в цитоплазме клеток, хотя синтез ДНК вируса осуществляется в ядре. Вирус имеет собственный транскрипционный фермент, ДНК-зависимую РНК-полимеразу.
Двухцепочечная ДНК, варьирующая по размеру от 170-190 п.о. в зависимости от изолята, сходна с ДНК вакцинного вируса, с ковалентно связанными концами и терминальными инвертированными повторами.
Исследования генома вируса показали, что он состоит из более консервативной центральной области (с вариабельным участком) размером около 125 т.п.о. и двух вариабельных областейпо концам вирусной ДНК, эти две вариабельные области генома наиболее сильно варьируют у различных изолятов вируса. Последние могут быть разделены на несколько групп в зависимости от нуклеотидного состава центрального участка генома.
| физико-химические свойства вируса АЧС |
| Он инактивируется при pH <4 и >11, хотя в зависимости от используемой питательной среды, на его инактивацию может уйти от нескольких часов до нескольких дней. |
| Он инактивируется при температуре 56 ºC в течение 60-70 мин и при 60 ºC в течение 20 мин. |
| Инактиванты/дезинфектанты: |
| 2% гидроксид натрия – 30 мин. |
| Гипохлорит (2-3% хлорин) – 30 мин. |
| Формалин (3/1000) – 30 мин. |
| Фенольные соединения |
| Йодистые вещества |
Геном изолятв Ba71V был полностью секвенирован. Его ДНК состоит из 170,701 нуклеотидов и содержит 151 открытую рамку считывания и 5 мультигенных семейств (МГС).
МГС локализованы раядом с терминальными инвертированными повторами в концевых участках генома. Существует как минимум 5 МГС, МГС360, МГС530, МГС10, МГС300, МГС505, и их состав наиболее разнообразен у различных изолятов вируса АЧС, что может свидетельствовать о том, что эти участки отвечают за антигенную вариабельность вируса, и возможный механизм уклонения вируса от иммунной системы организма-хозяина.
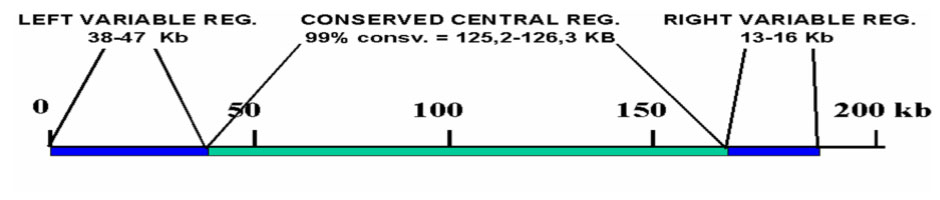
Структура генома вируса АЧС
| Генотипа вируса АЧС |
| Вирус АЧС не вызывает развития вируснейтрализующих антител, в следсвие чего не возможно серологическая классификация вируса. |
| Различия между изолятами определяют методы генотипирования вируса, основанные на секвенировании участка p72 гена B646L, разделяющего их на 22 генотипа |
| Субтипирование изолятов проводят при помощи секвенирования полного гена p54. Дополнительного разделения 22 генотипов вирус можно достичь секвенированием центрального вариабельного участка внутри гена B602L. |
| В Африке присутствуют все 22 генотипа вируса. Изоляты I генотипа в основном распространены на территории Западной Африки. Остальные 21 генотипы обнаруживают в восточных и южных регионах. |
| Прошлые вспышки АЧС в странах Европы, центральной и Южной Америки были вызваны I генотипом вируса. Однако, начиная с 2007 г, на Европейском континенте, Кавказском регионе, появился новый II генотип вируса, сходный с изолятами вируса циркулирующими на территории восточноафриканских стран. |
Всего 34 структурных белка и около 100 инфекционных протеинов ыло идентифицировано в макрофагах зараженных вирусом АЧС. Не менее 50 из них являются иммуногенными и вызывают выработку специфических антител при инфекции. Некоторые из этих белков p72, p54 (25 kda) и p12 являются структурными. Они участвуют в прикреплении вируса к клетке и являются высокоантигенными. Или как p32 участвуют в проникновении вируса в клетку. Тем не менее данные белки не связаны с индукцией иммунного ответа при болезни, но тем не менее они являются весьма полезными для для серологической диагностики.
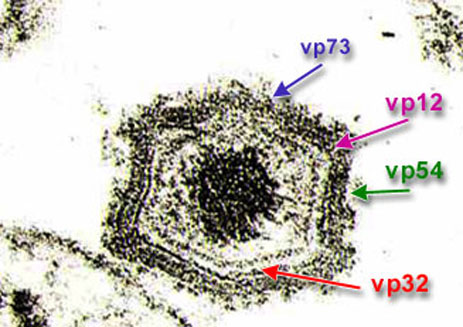
Расположение белков vp72, 32,
12 и 54 на вирусной частице.
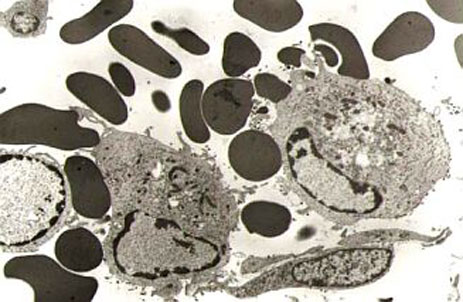
Вирус АЧС поражает
моноциты/макрофаги. Репродукция вируса
сопровождается развитием
феномена гемадсорбции.
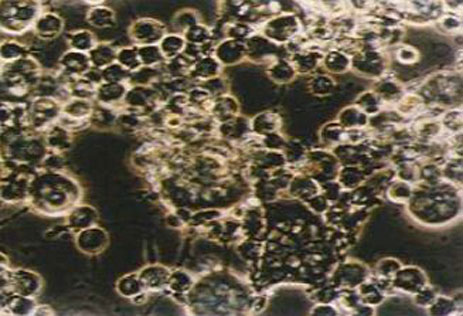
Цитопатический эффект, вызванный
вирусом АЧС в культуре свиных макрофагов
Вирус поражает моноциты/макрофаги иммунной системы организма животного, продуцирую развитие феномена гемадсорбции перед полным разрушением клетки. Вирусная репликация также наблюдается в эндотелиальных клетках, гепатоцитах, эпителии почечных канальцев и нейтрофилах. Репликация вируса также описана в Т- и В- лимфоцитах.
Вирус также может реплицироваться в естественных условиях в организме клещей O. Moubata и O. Erraticus.
Вирус АЧС адаптирован к росту на различных перевиваемых клеточных линиях с развитием цитопатического эффекта. Среди них для диагностических исследований и производства вирусных антигенов используют клеточные линии, полученные из почек обезьян МS и Vero.
Вирус попадает в клетку путем рецептор-опосредованного эндоцитоза.
Роль факторов гуморального и клеточного иммунного ответа в ходе развития АЧС до конца не известна. Сейчас описаны механизмы развития гуморального ответа и эффекторных клеток при АЧС, но до сих пор ничего не известно о их роли в защите против инфекции
Вирус АЧС вызывает развитие колоссального количества специфических антител, которые легко обнаруживаются лабораторными тестами начиная с седьмого дня после инфекции. Специфические антитела к вирусу АЧС присутствуют в организме инфицированного животного в течение нескольких лет после заражения и частично ответственны за отсрочку в появлении клинических признаков болезни и уменьшению виремии. Однако, антитела не способны полностью нейтрализовать вирус АЧС.