
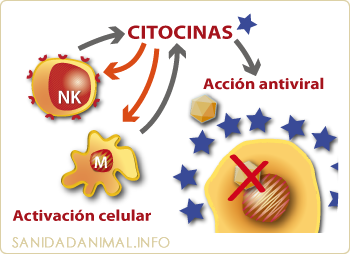
Si un antígeno atraviesa las barreras físicas y químicas se pondrá en marcha la inmunidad natural. La inmunidad natural es la primera barrera inmunológica no especifica del cerdo. En este tipo de respuesta, las citoquinas juegan un papel de gran importancia tanto de forma directa, por ejemplo, evitando la replicación viral gracias al interferón (IFN), como mediante diversos mecanismos inmunoreguladores que actúan, entre otras formas, como iniciadores de la inflamación, elevando la temperatura corporal, activando las células NK y los macrófagos. Las citoquinas que actúan en esta fase son fundamentalmente producidas por los monocitos-macrófagos y por otras células no inmunes como los fibroblastos y las células endoteliales.
Las citoquinas juegan un papel fundamental en la respuesta natural mediante mecanismos de acción directa frente al agente invasor (evitando la infección de las células por diferentes virus) o mediante mecanismos de activación celular (NK y macrófagos) que a su vez liberan más citoquinas
Las principales citoquinas que intervienen en la respuesta natural son:
| IL 1 |
| IL 6 |
| IL 12 |
| IL 16 |
| FACTOR DE NECROSIS TUMORAL (TNF α) |
| INTERFERÓN α y γ |
Las IL 1, IL 6, IL 12 intervienen en la respuesta natural activando a los monocitos-macrófagos y a las células NK, así como, facilitando la activación de los mecanismos de elevación de la temperatura corporal, para activar el sistema inmune y reducir el poder de multiplicación / replicación del agente patógeno.
El factor de necrosis tumoral (TNF α) es el principal iniciador de la respuesta inflamatoria, actúa sobre los vasos de la zona afectada aumentado la permeabilidad vascular, facilitando la extravasación a la zona de inmunoglobulinas, complemento, factores quimiotácticos, etc. De esta manera ayuda a en la defensa contra la agresión aportando mayores medios.
La citocina IL 16 parece estar ligada a la activación de linfocitos T CD 4+
CARACTERÍSTICAS DE LAS CITOQUINAS RELACIONADAS EN LA RESPUESTA NATURAL
| CITOQUINA | TIPOS | C. PRODUCTORA | FUNCIÓN |
| IL 1 | IL 1α IL 1β |
Macrófagos Linfocitos T y B Linfocitos NK |
Regulación inmunológica. Activación de Linfocitos T y Macrófagos |
| IL 2 | IL 2α IL 2β |
Linfocitos T (CD4) Linfocitos B y NK |
Activación Linfocitos T Activación Macrófagos |
| IL 4 | Linfocitos T activados | Th0 - th2 Expresión de SLA |
|
| IL 6 | IL6α. IL6β | Linfocitos Macrófagos Fibroblastos |
Activación linfocitos T y B Proinflamatoria |
| IL 10 | Linfocitos T y B Monocitos |
Diferenciación de linfocitos Activación Linfocitos T Secrección de Inmunoglobulinas |
|
| IL 12 | Neutrófilos Macrófagos Linfocitos T y B |
Diferenciación de Linfocitos (Th0 Th1 Th2) Activación de células NK |
|
| IL 16 | Linfocitos T | Quimiotaxis | |
| TNF α | TNFαR1 TNFαR2 |
Macrófagos Linfocitos CD4 Fibroblastos Mastocitos |
Inflamación local Cambios de permeabilidad Incremento fagocitosis |
| IFN α | Linfocitos | Activación SLA I | |
| IFN β | Fibroblastos Otras células |
Activación NK Inhibición de la replicación de los virus |
|
| IFN γ | Linfocitos T Células NK |
Inducción de SLA I y SLA II Activación macrófagos |

Resistencia transitoria
El interferón (IFN) que presenta los tipos: α, β y γ puede actuar de formas distintas para evitar la infección viral. La primera de ellas, inducidas por el IFN α y β, que son producidos fundamentalmente por monocitos-macrófagos y en menor proporción por fibroblastos, es la de inducir en las células susceptibles a la infección por virus un estado de resistencia transitoria frente a la infección de una gran gama de agentes víricos. Este efecto, de gran poder anti viral, no requiere de grandes cantidades de interferón y constituye uno de los mecanismos principales en la respuesta natural.
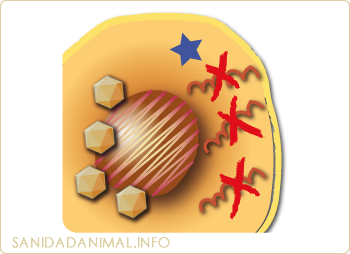
Degradación del ARN mensajero. Inhibición de la síntesis de proteína
El IFN también actúa mediante la síntesis de moléculas con actividad anti vírica, como la oligoadenilatosintetasa, que interfiere con la replicación del ADN y ARN o inhibiendo la síntesis de proteínas celulares, gracias a las distintas enzimas que producen las cuales, en presencia de dobles cadenas de ARN, inhiben la síntesis proteica mediante la activación de la RNAsa, la cual a su vez, degrada el ARN mensajero.
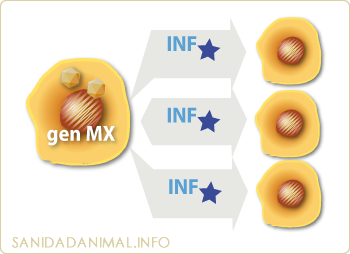
Activación de genes que expresan proteínas antivirales
Por último, los IFN α y β parecen activar diversos genes que expresan proteínas antivirales, como el gen Mx que determina hace resisten a células susceptibles contra la influenza murina. En este mecanismo, las células infectadas son capaces de secretar interferón que protege a las células vecinas, no-infectadas, de la infección.
Por otra parte, estos interferones aumentan la expresión del SLA I en las células infectadas, lo que favorece el reconocimiento de las mismas por los linfocitos CD 8+ y células NK con el consiguiente aumento de la actividad citotóxica

Incremento de la expresión del SLA
El IFN γ presenta una estructura distinta a los IFN α y β.
Es producido por los linfocitos T CD4+ y CD8+ y por las células NK tras recibir una estimulación antigénica. El IFN g, además de presentar actividad antiviral, interviene en muchas funciones inmureguladoras, tales como: Aumentar la expresión del SLA I (favorece la citotoxicidad) y del SLA II (favorece la colaboración celular para la presentación de los antígenos y la producción de anticuerpos)
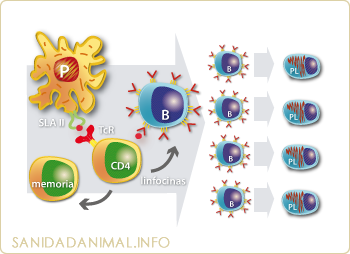
Cooperación celular para la presentación de antígenos y producción de anticuerpos. Las citoquinas juegan un papel fundamental en estos procesos |

Mediante las técnicas de citotoxicidad por liberación de cromo 51 se pueden valorar los mecanismos citotoxicos "in vitro". |
En el cerdo se ha detectado interferón durante la infección con varios virus (Gastroenteritis transmisible, Influenza, Peste porcina clásica, Enfermedad de Aujeszky), comprobándose, en algunas de ellas, su capacidad para aumentar o modular la expresión de los SLA.
El IFN-γ es una de las citoquinas que ha sido clonada en mas especies, y se han hecho gran cantidad de estudios de homologías y reactividad cruzada. De éstos estudios se concluye que estructuralmente se pueden clasificar en tres grandes grupos, según su homología funcional y estructural: El primer grupo estaría formado por el IFN γ humano y de otros primates, el segundo por los: bovinos, ovinos, ciervo, cerdos y caballos. En el tercero grupo estaría el ratón y la rata. En general, la mayoría de las citoquinas clonadas presentan una homología en cuanto a secuencia y por tanto de reactividad cruzada entre ellas.