
El grupo de estos genes se denominan: Complejo principal de histocompatibilidad, de la traducción de los términos ingleses "Mayor Histocopatibility Complex (MHC)". Estos genes juega un papel fundamental, no sólo en la respuesta inmune frente a los injertos o transplantes, sino también en el control de la presentación antigénica y en el desarrollo de la respuesta inmune. Las moléculas que codifican los genes MHC se unen a las proteínas extrañas para el animal para marcarlas y permitir que el sistema inmune las pueda reconocer y actuar frente a ellas.
DISTRIBUCIÓN DEL SLA PORCINO.
SLA I: Todas las células con núcleo.
SLA II: Monocitos, macrófagos, células dendríticas, linfocitos B
SLA III: No está en las células
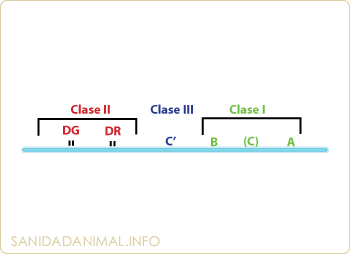
Todas las células porcinas, excepto los eritrocitos, presentan en su superficie unas proteínas, cuyo polimorfismo determina que exista o no rechazo entre los órganos transplantados de dos animales diferentes. Estas proteínas del Complejo principal de histocompatibilidad (MHC), en el cerdo, se denominan SLA, siglas que provienen de las palabras inglesas "Swine Leucocyte Antigens". Se describieron por vez primera a principios de los años 70, al observarse una correlación entre el rechazo agudo en el transplante y un grupo de antígenos expresados en la superficie de los linfocitos periféricos. Los genes responsables de la codificación de los SLA se encuentran localizados en el cromosoma 7 con una dimensión aproximada de 2 Mb, de los que alrededor de 70 genes han sido ya caracterizados. Están ligados a los genes que codifican para los sistemas que J y C. Al igual que en otras especies animales, en el cerdo se han descrito tres antígenos de histocompatibilidad en función a su estructura química, su distribución tisular y su función, denominados como:
| SLA I | SLA II | SLA III |
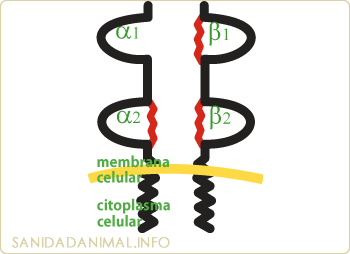
Los antígenos de clase I y II son proteínas integradas en la membrana celular. Su estructura la forman: Una región extracelular, un segmento que atraviesa la membrana y una pequeña parte que penetra en el citoplasma.
Los antígenos de la clase I y III están separados por el centrómero del cromosoma 7 de los de clase II, presentando una relación espacial semejante a los MHC humanos.
Los antígenos de clase I se expresan en la superficie de todas las células con núcleo con la única excepción de las neuronas y los trofoblastos. Según se van formando en la célula infectada (infección viral), se enlazan con las proteínas virales (antígenos) que se están sintetizando, formando un complejo SLA I - Antígeno que se expresa en la membrana de la célula infectada. Este complejo será reconocido por un determinado linfocito T, el CD 8+ o linfocito T citotóxico, que destruirá la célula infectada. La presentación de antígenos en células infectadas a los CD8+, es una de sus funciones principales de los SLA I.
Los antígenos de la clase I están formados por un heterodímero compuesto por dos cadenas: una pesada denominada a de 45 kd de peso molecular y extremadamente polimórfica y codificada por genes del SLA. La otra cadena es ligera de peso molecular inferior (12 kb) denominada B, no está codificada por genes SLA y no es polimórfica. Cada haplotipo del SLA codifica 2 o 3 loci clase I, denominados, al igual que en la especie humana, como : A, B y C con un total de entre 7 a 10 genes diferentes. Las diferencias funcionales de los genes clase I se pueden definir serológicamente, habiéndose identificado más de 40 alelos diferentes del SLA I
Los antígenos de clase II se expresan, en el cerdo, de forma más restringida. Se encuentran en los linfocitos B, las células presentadoras de antígenos y en varias subpoblaciones de linfocitos T, estos últimos tanto si están activados como sin estarlo (particularidad de la especie porcina). La función del SLA II es también la presentación de antígenos a los linfocitos T CD4+, pero en este caso, mediante las células fagocíticas o presentadoras de antígenos, las cuales procesan los antígenos por degradación mediante enzimas y no por infección celular como en el caso de los SLA I. Las moléculas del agente capturado por estas células y degradadas en pequeñas partículas, son asociadas a los SLA y expresadas en la membrana de la célula formando un complejo: SLA II - Antígeno. En este caso el linfocito T que reconocerá al mencionado complejo será el CD4 o linfocito T cooperador. También el linfocito B expresa SLA II. (capítulo 3)
Genes del SLA II
| 1 SLA-DRA |
| 2 SLA-DRB |
| 1 o 2 SLA-DQA |
| 2 o 3 SLA-DQB |
| 1 SLA-DPA |
| 1 SLA-DPB |
Los antígenos de clase II están representados por los loci: SLA-DR y SLA-DQ. Los antígenos de clase II están formados por dos cadenas de glicoproteínas denominadas α y β que presenta un peso molecular de: 33 a 35 kb la cadena α y de 27 a 29 la cadena β. Ambas cadenas están codificadas por genes del SLA α y β que presenta un peso molecular de: 33 a 35 kb la cadena alfa y de 27 a 29 la cadena B. La familia SLA está formada por alrededor de 10 genes diferentes.
El SLA III codifica:
| Factores de complemento: C4 y C2 |
| 21-Hidroxilasa |
| Factor de Necrosis Tumoral |
Los genes de clase III: A diferencia de los SLA I y SLA II, codifican proteínas que no se encuentran en la superficie de las células sino en la sangre. Así varios de los componentes del complemento son codificados por el SLA III, interviniendo además, en otras acciones del sistema inmune menos especificas, como la selección de factores de necrosis tumoral.
Los antígenos del SLA regulan la respuesta inmune frente a un gran número de agentes patógenos, jugando un papel fundamental en los mecanismos de presentación antigénica.
EL RECONOCIMIENTO DE LOS ANTÍGENOS POR LOS LINFOCITOS SE REALIZA POR LA ASOCIACIÓN A LOS SLA.