|
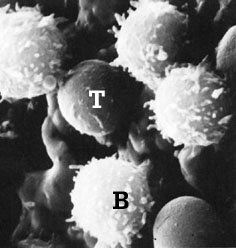
Image of T and B lymphocytes obtained using a scanning electron microscope
|
|
From a morphological point of view, differences between T and B lymphocytes are not found when an optical microscope is used. Both lymphocytes are cells measuring 7 to 9 microns, with a voluminous nucleus and limited cytoplasm. Notable differences can be found when a scanning electron microscope is used. The T lymphocytes have a smooth and even surface, whilst the B lymphocytes have multiple projections which correspond to surface immunoglobulins. |
|
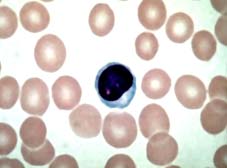
Image of a lymphocyte stained with Giemsa, observed by optical microscopy
|
Over recent decades, different surface markers have been designed to differentiate the different populations of lymphocytes. However the development of monoclonal antibodies (MA) produced against the different porcine lymphatic cells have enabled the differentiation of these populations, and their grouping according to their membrane antigens and their function. |
| The MAbs have enabled a third lymphoid population to be defined, known as null cells as they do not have the traditional markers found in T and B lymphocytes (rosette formation with sheep erythrocytes and surface immunoglobulins, respectively). These null cells, which in pigs can represent between 9 and 19% of circulating lymphocytes, are actually a subpopulation of T lymphocytes (T lymphocytes g-d) |
|
|
Main receptors traditionally used to differentiate between T and B lymphocytes |
| B lymphocytes: |
|
| T lymphocytes: |
Rosettes with sheep erythrocytes. Esterase-positive. |
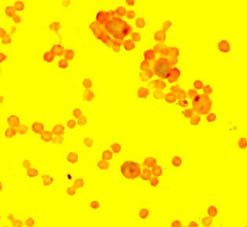
Porcine T lymphocytes forming rosettes with sheep erythrocytes.
|
|
|
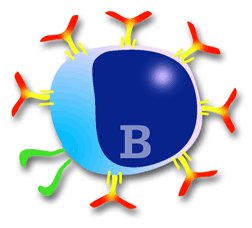
Diagram of a B lymphocyte
|
B lymphocytes in pigs are made in the bone marrow at a proportion of between two-hundred and four-hundred million per day, demonstrating the enormous capacity for response of the immune system. . Remember that a B lymphocyte produces a specific antibody (1 cell = 1 type of antibody). In the peripheral blood, B lymphocytes make up 8-18% of total lymphocytes in pigs. The membrane of B lymphocytes is made of many molecules, many of which have been studied thanks to the MAbs. Amongst these, one of the most important is the BcR (B Cell Receptor) complex. |
|
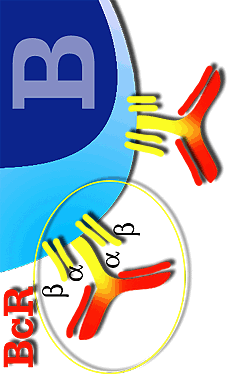
Details of the B lymphocyte BcR complex
|
The BcR consists of several chains. Some are variable (they are immunoglobulins), in which each B lymphocyte presents different variations according to the type of immunoglobulin (mainly IgM and IgG) or (IgA and IgE) or according to the type of antigen. The other two chains are invariable (formed by two chains a and b), and common to all B lymphocytes. The job of the variable chains, which are immunoglobulins, is to react with the specific antigen, whilst the invariable chains serve to transmit the signal to the interior of the cell to start antibody production.
The B lymphocytes are characterized by the presence of immunoglobulins on their surface, mainly of the type IgM and IgG, allowing them to react with antigen in its native form (the T lymphocytes cannot react with the antigen in its native form) (Chapter 3). Most antigens that react with B lymphocytes are protein in nature, although they can also react with polysaccharides. The initiation of the reaction with the antigen is always carried out through the membrane immunoglobulins (BcR signal), but the production of antibodies needs the collaboration of CD 4+ lymphocytes in most cases. (chapter 3). Antigens which need this cooperation with CD 4 are known as: T dependent antigens (these are the vast majority). Other antigens such as some lipopolysaccharides or bacterial polysaccharides do not need T lymphocytes for the stimulation and production of antibodies. This type of antigen is known as: T independent antigens. |
|
Other markers of B lymphocytes have been differentiated through the MAbs. These markers have been grouped into "cluster of differentiation" (CD), based on their antigenic and functional differences.
|
MAIN CD DEVELOPED FOR THE STUDY OF B LYMPHOCYTES |
| CD1: The CD1 antigen, which is very similar to the SLA I, can be observed in B lymphocytes (sIg+) in peripheral blood in a proportion of 30 to 60% and in T lymphocytes of the thymus cortical zone in around 60% of the total. |
| CD 21: Observed in the membrane of mature B lymphocytes. |
| CD 45: Observed in B lymphocytes and in some subpopulations of T lymphocytes. |
| SWC7: Found in the B lymphocytes situated in the lymphoid organs, this is not found in the lymphocytes of peripheral blood. |
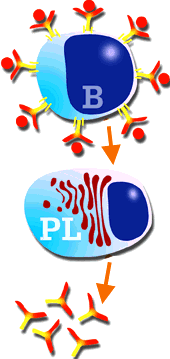
Diagram of the stimulation of a B lymphocyte, transformation in plasma cell and the production of antibodies.
|
A B lymphocyte stimulated by either a T-dependent or T-independent antigen, generates a plasma cell clone which will produce and secrete a large quantity of antibodies. These antibodies are specific against the epitope which induced the immune response. This immune response, mediated only by antibodies is known as: humoral response. The stimulated B lymphocyte or plasma cell has a different morphology to the original B lymphocyte (large nucleus and small cytoplasm) and more typical of a "factory cell" with a small nucleus and a large cytoplasm. These cells are mainly located in the lymph nodes, red pulp of the spleen, bone marrow, and intestinal and respiratory mucous membranes. "In vivo" their average life is very short, of only 2 to 3 days, with "in vitro" even shorter at only a matter of hours. |
|
|
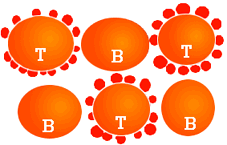
Diagram showing the formation of rosettes in pig T lymphocytes with sheep erythrocytes.
|
Pig T lymphocytes are produced in the thymus and in smaller amounts in the T-dependent zones of the secondary lymphoid organs. They differ from B lymphocytes in that they do not present immunoglobulins on their surface, but form rosettes with sheep erythrocytes. The T lymphocytes play a key role in the immune response, as they favour the activation of mechanisms of presentation of antigens to B lymphocytes for the production of antibodies and are also responsible for cell immunity (immune response mediated by cells rather than antibodies).
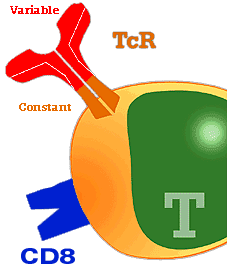
As well as SLA, two further types of antigen specific receptors are found in the membrane of the lymphocytes. These are named T cell receptors or TcR. These receptors are:
Receptor a-b (TcR a-b). They represent between 40 to 60% of the lymphocytes found in peripheral blood. The a b LYMPHOCYTES are: CD4+ helper lymphocytes and CD 8+ cytolytic lymphocytes
Receptor g- d (TcR g-d). Present in most of the cells formerly known as null cells. They are now known as g T LYMPHOCYTES |
|
|
|
These receptors are made of two heavy and two light chains, similar to immunoglobulin chains. Their function is to react with antigens which are presented by antigen-presenting cells and cells associated with SLA.
As is the case with the B lymphocytes, thanks to the production of AcM against porcine lymphocytes, several subpopulations of pig T lymphocytes have been grouped into "cluster of differentiation" molecules (CD) based on their antigenic and functional differences.
Thanks tos it has been proved that the pig presents certain differences to other animal species and with humans. In pigs there is a lymphocyte population which is double positive to CD4+ and CD8+ (CD4+CD8+) which increases in proportion to the age of the animal. Therefore, when the animal is one week old the percentage of CD4+CD8+ represents less than 2% of lymphocytes and at 3 years old it represents around 30%. The function of these double positive lymphocytes is believed to be related to the memory cells, although recently it has been related to the helper activity of the T lymphocytes in primary infections. It thus seems that there are two populations of helper cells, CD4+CD8- and CD4+CD8+. Both populations act in cell cooperation in the primary response, at least when "in vitro", whilst in the secondary response only the CD4+CD8+ act. It is known that as well as increasing with age, these double positive cell populations are found in significant amounts in the tonsils (50%) and the lymph nodes (30%) of adult animals. In regard to cytotoxic lymphocytes, two subpopulations of CD4-CD8+ can be differentiated by virtue of CD 6 expression. Therefore, lymphocytes that express antigen CD 6- are related to spontaneous cytotoxicity whilst CD 6+ SLA I are related to cytotoxicity of virus-infected cells.
|
MAIN CD DEVELOPED FOR THE STUDY OF T LYMPHOCYTES |
|
CD 1: Found in 60% of T lymphocytes of the thymus cortex. They are very similar to SLA I. |
|
CD 2: TcR re a-b (T a b). The T lymphocytes with TcR gd (T LYMPHOCYTES g d) are either negative to CD2- or express it in very low percentages (3 to 6%). These lymphocytes (g- d), formerly known as null cells, are identified by the monoclonal antibody SWC6. |
|
CD 3: Present in lymphocytes related to cellular activation and suppression. They are of great importance in xenotransplants. |
|
CD 4: Present in T helper cells and in those responsible for delayed hypersensitivity reactions. CD4+ lymphocytes recognise SLA II. |
|
CD 6: Present in cells related to cytotoxicity. |
|
CD 8: Located in cytotoxic T lymphocytes. CD 8+ lymphocytes recognise SLA I. |
|
CD 16: To differentiate natural killer cells. |
|
|
Lymphocytes Th1 and Th2 When lymphocyte CD4 is in the proliferation phase, after antigen presentation (chapter 3) the CD 4 lymphocytes pass through a differentiation phase known as Th0 which is subsequently differentiated both in its actions and production of cytokines (chapter 6), favouring the cell response (Th1 activation) or the humoral response (Th2 activation). The change from Th0 to Th1 or to Th2 depends on the signal received by Th0 during antigen presentation. The mechanisms by which one or another signal is produced is still little understood but seems to depend on the nature of the pathogen (Th1 in intracellular), of the cytokines liberated by the presenting-cell, of the antigen dose, etc.
The Th1 lymphocytes basically stimulate the response of the CD8 lymphocytes, whilst the Th2 lymphocytes are more specialized in the activation of B lymphocytes for the production de antibodies.
Finally, another peculiarity of swine T lymphocytes is that they express SLA II in their membranes. Compared to humans or mice, whose T lymphocytes do not have class II histocompatibility antigen when not activated. |
| |