
El tejido linfoide asociado a las mucosas (MALT) forma parte del sistema inmune aunque, con cierta independencia del sistema sistémico. Es el encargado de proteger las mucosas del cerdo del ataque de los agentes patógenos, tanto en una respuesta primaria como secundaria. Está formado por nódulos de tejido linfoide que, según su localización, se denominan: GALT y BALT
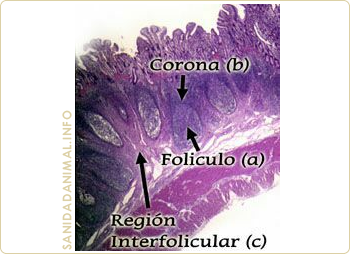
La denominación GALT proviene de las palabras inglesas "Gut Associated Lymphoid Tissues" y cuya traducción sería: Tejido linfoide asociado al intestino. El GALT está formado por todo el tejido linfoide que se encuentra en las paredes intestinales (ganglios, placas de Peyer, folículos linfoides aislados). (órganos linfoides secundarios)
La denominación BALT tienen su origen en las palabras inglesas "Broncus Associated Lymphoid Tissues" en español: tejido linfoide asociado a los bronquios. Está formado por todo el tejido linfoide (tonsilas, ganglios, folículos linfoides) localizado en las mucosas respiratorias, desde las fosas nasales hasta los pulmones.
Una particularidad del BALT en la especie porcina, a diferencia de los roedores o de la especie humana, es la gran presencia, en los pulmones, de macrófagos intravasculares que presentan gran actividad.
Una ejemplo de la gran importancia, que el tejido linfoide de las mucosas presenta en los mecanismos de defensa del cerdo frente a las infecciones, lo prueba la gran cantidad de tejido linfoide disponible.
Este tejido linfoide se distribuye de forma estratégica en las siguientes zonas:
- Zonas de procesamiento e inicio de la respuesta inmune. Zonas Inductoras.
- Zonas de respuesta (humoral y celular) denominadas. Zonas efectoras.
Las zonas de inicio o inductoras de la respuesta inmune de las mucosas, disponen de elementos semejantes a los componentes del sistema inmune sistémico para realizar la captación de los antígenos e iniciar la respuesta inmune. Con la única diferencia de las células M, que son unas células epiteliales especializadas en el transporte de antígenos, los demás componentes (células presentadoras, linfocitos T y linfocitos B) actúan de forma similar al sistema sistémico. Estos componentes celulares están localizados en: las tonsilas, placas de Peyer, ganglios y en el tejido linfoide difuso. En definitiva, tanto en las zonas GALT como las BALT (órganos secundarios del cerdo) tiene lugar el contacto con el antígeno, transporte, procesamiento y presentación a los linfocitos T y B.
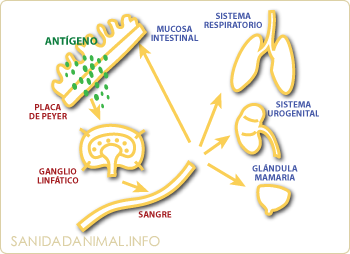
Estimulación del tejido linfoide de las mucosas BALT o GALT. Este mecanismo permite que, aunque la estimulación antigénica haya sido local, la respuesta inmune sea generalizada.
Los antígenos suelen entrar en el animal por inhalación o ingestión. Mediante un proceso ligado a las células M, CPAg o directamente sobre linfocitos B (semejante al descrito en cooperación celular), habrá una estimulación de células B (precursoras fundamentalmente de IgA) y de linfocitos T. Las células estimuladas en estas zonas de inicio, abandonan el área mediante el sistema sanguíneo, migrando hacia las diferentes zonas efectoras.
Este mecanismo permite, que aunque la estimulación antigénica haya sido a nivel local, la respuesta inmune sea generalizada en todo el organismo del animal, por lo que se define como: respuesta inmune secretora y generalizada.

Esquema de la estructura de una inmunoglobulina IgA
En las zonas efectoras la mayor parte de las células inmunes son linfocitos T, que se encuentran entre las células epiteliales (linfocitos intraepiteliales) y por debajo de ellas en la lamina propia. Fundamentalmente CD 8 (77%) y CD 4 γ-δ. También hay linfocitos B, que pueden reaccionar con el antígeno.
Las células plasmáticas productoras, fundamentalmente de inmunoglobulina del isotipo IgA, se encuentran principalmente en los ganglios linfáticos y en las células plasmáticas difusas, que se localizan en las paredes del intestino y sistema respiratorio. Estas células son fundamentales en la respuesta inmune de las mucosas, secretando alrededor del 80% de IgA, con la única excepción de las tonsilas, donde la inmunoglobulina sintetizada de forma mayoritaria es la IgG seguida de la IgA.
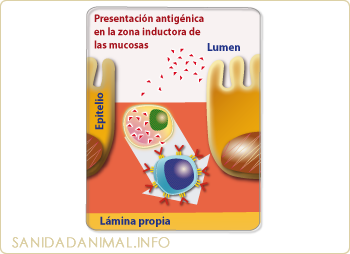
PRESENTACIÓN ANTIGÉNICA EN LA ZONA INDUCTORA DE LAS MUCOSAS. El antígeno que penetra en los enterocitosis se destruye rápidamente por los lisosomas. Sin embargo, el antígeno que es capturado por las células M, es trasportado sin degradación y presentado a los linfocitos intraepiteliales. A partir de ahí podrá pasar a los ganglios linfáticos
El transporte de los antígenos a las zonas inductoras (Placas de Peyer y folículos linfoides) se realiza fundamentalmente mediante las células M. Las células M, son células epiteliales especializadas en el transporte de antígenos. No actúan enzimáticamente sobre los antígenos. Las células M captan a los antígenos de la luz del intestino y los transportan completamente intactos a la presentación de los linfocitos intraepiteliales (dentro de la propia célula) o pasan por el espacio intercelular hacia el líquido hístico y presentan el antígeno a las células presentadoras (macrófagos, células dendríticas y linfocitos B) presentes en el espacio subepitelial o en la lamina propia. Los mecanismos de activación a nivel de la lamina propia siguen un esquema semejante al descrito en la cooperación celular.
Esquema de la estructura de una inmunoglobulina IgA
En las zonas efectoras la mayor parte de las células inmunes son linfocitos T, que se encuentran entre las células epiteliales (linfocitos intraepiteliales) y por debajo de ellas en la lamina propia. Fundamentalmente CD 8 (77%) y CD 4 γ-δ. También hay linfocitos B, que pueden reaccionar con el antígeno.
Las células plasmáticas productoras, fundamentalmente de inmunoglobulina del isotipo IgA, se encuentran principalmente en los ganglios linfáticos y en las células plasmáticas difusas, que se localizan en las paredes del intestino y sistema respiratorio. Estas células son fundamentales en la respuesta inmune de las mucosas, secretando alrededor del 80% de IgA, con la única excepción de las tonsilas, donde la inmunoglobulina sintetizada de forma mayoritaria es la IgG seguida de la IgA.
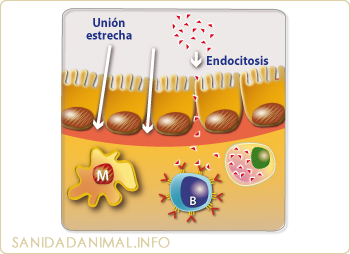
El antígeno en la zona efectora, entra por endocitosis o a través de las uniones estrechas.
En las zonas efectoras también pueden presentarse antígenos, aunque el mecanismo de entrada suele ser diferente al de las zonas inductoras. En las zonas efectoras el antígeno puede acceder por endocitosis mediante las células epiteliales o atravesando por las llamadas zonas o uniones estrechas. La captura y presentación se realiza mediante macrófagos, células M o linfocitos B y los pasos siguientes siguen un procedimiento al anteriormente descrito.
La capacidad de inducción de una respuesta inmune a nivel de las mucosas, generalmente requiere de mayor cantidad de antígeno y a veces mayor número de inmunizaciones que en el sistema sistémico, sobre todo en las inmunizaciones por vía oral.
Esto es debido a las múltiples alteraciones y degradaciones enzimáticas que sufren los antígenos por esta vía. Este mecanismo es bueno para la defensa del animal, pero hay que tenerlo en cuenta a la hora de diseñar vacunas orales. Aunque como ya veremos en el capítulo 8, existen varias estrategias para inducir una buena respuesta inmune también por vía oral. No obstante, es generalmente más fácil, inducir inmunidad en las mucosas respiratorias mediante una inmunización oral, que inducir respuesta inmune en las mucosas intestinales, mediante inmunización nasal.
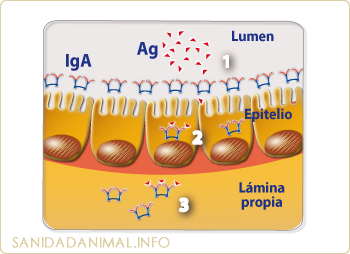
ESQUEMA DE ACTUACIÓN DE LA IgA. La IgA puede actual a nivel de la luz intestinal (lumen), evitando la adherencia al epitelio de virus y/o bacterias (1), dentro de los enterocitos incluso neutralizando virus (2) y por último, en el líquido hístico (3).
La IgA (capítulo 4) juega un papel importantísimo en la respuesta inmune de las mucosas. Su configuración en forma de dímero o como tretrámero, le permite disponer de entre 4 a 8 sitios de unión al antígeno, lo que la hace tremendamente efectiva frente a diferentes antígenos bacterianos, mediante reacciones del tipo ADCC, ya que la IgA no es bactericida. Aunque si presenta gran capacidad para la neutralización de algunos virus, incluso dentro de las células epiteliales. Es la única inmunoglobulina capaz de poder actuar dentro de una célula, pero sobre todo su principal actividad en la defensa de las mucosas es la de evitar la adherencia de bacteria y virus a la superficie del epitelio. La IgA, por tanto puede actuar en tres lugares y de forma distinta. Por un lado, puede unirse al antígeno en la luz intestinal, para evitar la adherencia de virus y/o bacterias a la superficie del epitelio o dentro de los enterocitos, y por último, en el líquido hístico.
Una muestra más de la importancia que esta inmunoglobulina presenta en los mecanismos de defensa de la especie porcina, lo representa el hecho que en la cerda, el 85% de las células que contienen inmunoglobulinas en la lamina propia del intestino presentan IgA.