|
|||||
|
|
COME INTERVENGONO LE CITOCHINE NELLA RISPOSTA NATURALE O INNATA?
|
||||
|
|||||
|
|
COME INTERVENGONO LE CITOCHINE NELLA RISPOSTA NATURALE O INNATA?
|
||||
|
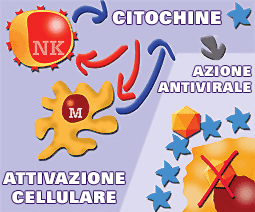
Le citochine hanno un ruolo fondamentale nella risposta naturale o innata attraverso meccanismi d'azione diretta contro l'agente invasore, nella fase precoce dell'infezione, o mediante meccanismi immunoregolatori che portano l'attivazione delle cellule NK e dei monociti-macrofagi con la conseguente liberazione di altre citochine.
|
||||||||||||||||||||||||||||||||||||||
|
Se un antigene attraversa le barriere fisiche e chimiche si metterà in moto l'immunità naturale. L'immunità naturale è la prima barriera immunologica non specifica del suino. In questo tipo di risposta, le citochine hanno un ruolo di grande importanza sia in forma diretta, ad esempio, evitando la replicazione virale grazie all'interferone (IFN), sia mediante diversi meccanismi immunoregolatori che agiscono, tra le altre forme, come iniziatori dell'infiammazione, elevando la temperatura corporea, attivando le cellule NK ed i macrofagi. Le citochine che agiscono in questa fase sono fondamentalmente prodotte dai monociti-macrofagi e da altre cellule non immuni come i fibroblasti e le cellule endoteliali.
|
Le citochine hanno un ruolo fondamentale nella risposta naturale mediante meccanismi d'azione diretta contro l'agente invasore (evitando l'infezione delle cellule da diversi virus) o mediante meccanismi d'attivazione cellulare (NK e macrofagi) che a loro volta liberano più citochine. |
|||||||||||||||||||||||||||||||||||||
|
Le principali citochine che intervengono nella risposta naturale sono:
|
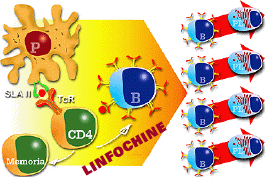
Le IL 1, IL 6, IL 12 intervengono nella risposta naturale attivando i monociti-macrofagi e le cellule NK, ed anche facilitando l'attivazione dei meccanismi di elevazione della temperatura corporea, per attivare il sistema immunitario e ridurre il potere di moltiplicazione / replicazione dell'agente patogeno.
La citochina IL 16 sembra essere legata all'attivazione dei linfociti T CD 4+. |
|||||||||||||||||||||||||||||||||||||
|
CARATTERISTICHE DELLE CITOCHINE RELAZIONATE NELLA RISPOSTA NATURALE
|
||||||||||||||||||||||||||||||||||||||
|
L'interferone (IFN) che presenta i tipi: a, b e g può agire in diversi modi per evitare l'infezione virale. Il primo metodo, indotto dall'IFN a e b, che sono prodotti fondamentalmente da monociti-macrofagi ed in minor proporzione da fibroblasti, è quello di indurre nelle cellule a rischio di infezione dal virus uno stato di resistenza transitoria contro l'infezione di una grande gamma di agenti virali. Questo effetto, anti-virale, non ha bisogno di grandi quantità di interferone e costituisce uno dei meccanismi principali nella risposta naturale. |
Resistenza transitoria |
|||||||||||||||||||||||||||||||||||||
|
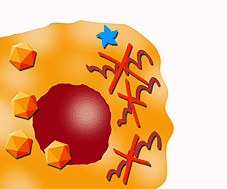
L'IFN agisce anche mediante la sintesi di molecole con attività anti-virale, come ad esempio la oligoadenilatosintetasi, che interferisce sulla replicazione del DNA e RNA o inibisce la sintesi delle proteine cellulari; o mediante diversi enzimi che in presenza di catene doppie di RNA inibiscono la sintesi proteica tramite l'attivazione di una RNAs, che è in grado di degradare il RNA messaggero. |
Degradazione del RNA messaggero. Inibizione della sintesi di proteine |
|||||||||||||||||||||||||||||||||||||
|
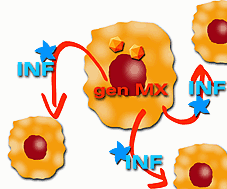
Per ultimo, gli IFN a e b pare che attivino diversi geni che esprimono proteine antivirali, come il gene Mx che determina in cellule sensibili resistenza contro l'influenza murina. In questo meccanismo, le cellule infette sono capaci di secretare interferone che protegge le cellule vicine, non-infette, all'infezione.
D'altra parte, questi interferoni aumentano l'espressione del SLA I nelle cellule infette, favorendo il riconoscimento delle stesse dai linfociti CD 8+ e cellule NK con il conseguente aumento dell'attività citotossica.
|
Attivazione dei geni che esprimono proteine antivirali |
|||||||||||||||||||||||||||||||||||||
|
Incremento dell'espressione del SLA |
L'IFN g presenta una struttura diversa dai IFN a e b. E' prodotto dai linfociti T CD4+ e CD8+ e dalle cellule NK dopo avere ricevuto una stimolazione antigenica. L'IFN g, oltre a presentare una attività antivirale, interviene in molte funzioni immunoregolatrici, come ad esempio: Aumentare l'espressione del SLA I (favorisce la citotossicità) e del SLA II (favorisce la collaborazione cellulare per la presentazione degli antigeni e la produzione di anticorpi) |
|||||||||||||||||||||||||||||||||||||
|
Cooperazione cellulare per la presentazione di antigeni e produzione di anticorpi. Le citochine hanno un ruolo fondamentale in questi processi |
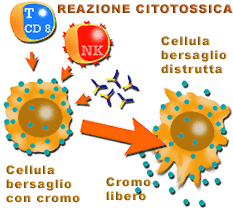
Mediante le tecniche di citotossicità per liberazione di cromo 51 si possono valutare i meccanismi citotossici "in vitro". |
|||||||||||||||||||||||||||||||||||||
|
Nel suino è stato rilevato l'interferone durante l'infezione con diversi virus (Gastroenterite trasmissibile, Influenza, Peste Suina Classica, Malattia di Aujeszky), ed è stato possibile verificare, in alcune di esse, la sua capacità di aumentare o modulare l'espressione dei SLA. |
||||||||||||||||||||||||||||||||||||||
|
L'IFN-g è una delle citochine clonate in diverse specie, e sono stati realizzati diversi studi di omologie e reattività incrociata. Da questi studi risulta che strutturalmente si possono classificare in tre grandi gruppi, secondo la sua omologia funzionale e strutturale: Il primo gruppo è formato dal IFN g umano e di altri primati, il secondo da: bovini, ovini, cervo, suini e cavalli. Nel terzo gruppo si trova il topo. In generale, la maggior parte delle citochine clonate presentano omologie in quanto a sequenza e di conseguenza reattività crociata tra di esse. |
||||||||||||||||||||||||||||||||||||||