|
|
RUOLO
DELLE MUCOSE. |
|
||||
|
|
RUOLO
DELLE MUCOSE. |
|
Le mucose hanno un ruolo molto importante nella difesa immunologica del suino, dato che un gran numero di agenti patogeni utilizzano le mucose come via di ingresso.
Il tessuto
linfatico associato alle mucose forma parte del sistema immunitario, anche se con una certa
indipendenza. E' il responsabile della protezione delle mucose del
suino dagli attacchi degli agenti patogeni, sia nella risposta primaria
sia in quella secondaria.
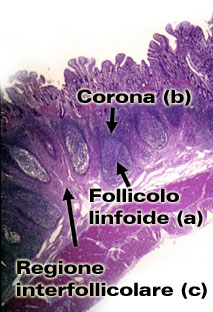
E' formato da noduli di tessuto linfatico che, secondo la loro
localizzazione, vengono denominati: GALT
e BALT |
||||||
|
La denominazione GALT proviene dall'inglese "Gut Associated Lymphoid Tissues" che tradotto significa tessuto linfatico associato all'intestino. Il GALT è formato dal tessuto linfatico che si trova nelle pareti intestinali (gangli, placche di Peyer, follicoli isolati). (organi linfatici secondari) La denominazione BALT proviene dall'inglese "Broncus Associated Lymphoid Tissues", che tradotto significa tessuto linfatico associato ai bronchi. E' formato dal tessuto linfatico (tonsille, gangli, follicoli linfatici) localizzato nelle mucose respiratorie dalle cavità nasali fino ai polmoni. |
|
|||||
|
Una particolarità del BALT nella specie suina, a differenza dei roditori o della specie umana, è la massicia presenza, a livello polmonare, di macrofagi intravascolari molto attivi. |
||||||
|
IMMUNITA' DELLE MUCOSE. |
||||||
|
La grande quantità di tessuto linfatico disponibile dimostra la notevole importanza delle mucose nei meccanismi di difesa del suino contro le infezioni. Questo tessuto linfatico si distribuisce in modo strategico nelle seguenti zone:
Le zone
di inizio o induttrici della risposta immunitaria delle mucose,
dispongono di elementi somiglianti ai componenti
del sistema immunitario sistemico per realizzare la captazione
degli antigeni ed iniziare la risposta immunitaria. Con
l'unica differenza delle cellule M, che
sono cellule epiteliali atte al trasporto di antigeni, gli
altri componenti (cellule presentatrici, linfociti
T e linfociti B) agiscono in modo simile al sistema generale. Questi
componenti cellulari sono localizzati in: tonsille, placche di Peyer,
gangli e tessuto linfatico diffuso. In definitiva, sia nelle
zone GALT che nelle BALT (organi
secondari del suino) avviene il contatto con l'antigene, il
trasporto, l'elaborazione e la presentazione dei linfociti T e B. |
||||||
|
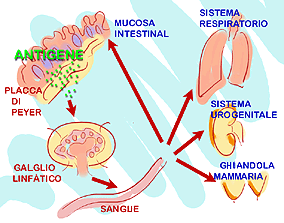
Questo meccanismo permette che, nonostante la stimolazione antigenica sia avvenuta a livello locale, la risposta immunitaria sia generalizzata in tutto l'organismo animale, per questo viene definita come: risposta immunitaria secretoria e generalizzata. |
|
|||||
|
Stimolazione del tessuto linfatico delle mucose BALT o GALT. Questo meccanismo permette che, con una stimolazione avvenuta a livello locale, la risposta immunitaria sia generalizzata. |
||||||
|
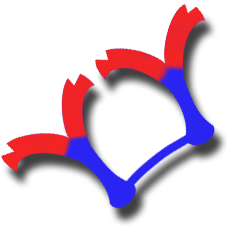
Schema della struttura di una immunoglobulina IgA |
Nelle zone effettrici la maggior parte delle cellule immunitarie sono linfociti T, che si trovano tra le cellule epiteliali (linfociti intraepiteliali) e sotto di esse nella lamina propria. Fondamentalmente CD 8 (77%) e CD 4 g-d. Ci sono anche linfociti B, che possono reagire con l'antigene. Le cellule plasmatiche produttrici, fondamentalmente di immunoglobulina dell'isotipo IgA, si trovano principalmente nei gangli linfatici e nelle cellule plasmatiche diffuse, che si localizzano nelle pareti dell'intestino e nel sistema respiratorio. Queste cellule sono fondamentali nella risposta immunitaria delle mucose, secernendo attorno all'80% di IgA, con l'unica eccezione delle tonsille, dove l'isotipo maggiormente sintetizzato è la IgG seguita dalla IgA. |
|||||
|
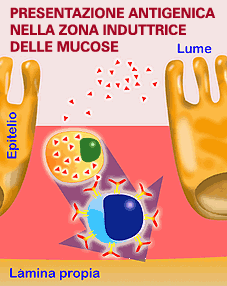
Il trasporto degli antigeni alle zone induttrici (Placche di Peyer e follicoli linfatici) si realizza fondamentalmente mediante le cellule M. Le cellule M, sono cellule epiteliali specializzate nel trasporto di antigeni. Non agiscono enzimaticamente sugli antigeni. Le cellule M captano gli antigeni del lume intestinale e li trasportano intatti ai linfociti intraepiteliali (all'interno della propria cellula) o passano attraverso lo spazio intercellulare verso il liquido tessutale e presentano l'antigene alle cellule presentatrici (macrofagi, cellule dendritiche e linfociti B) presenti nello spazio sub-epiteliale o nella lamina propria . I meccanismi di attivazione della lamina propria seguono uno schema simile a quello descritto nella cooperazione cellulare. |
L'antigene che penetra negli enerocitoti viene distrutto rapidamente dai lisosomi. Invece, l'antigene catturato dalle cellule M, è trasportato senza degradazione e presentato ai linfociti intraepiteliali. Da lì potrà passare ai gangli linfatici.
|
|||||
|
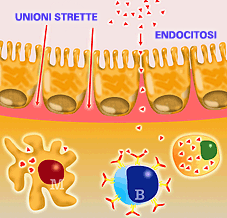
L'antigene
nel la zona effettrice, entra per endocitosi o attraverso le unioni
strette. |
Le zone effettrici possono anche presentare antigeni, tuttavia il meccanismo di entrata è di solito diverso rispetto a quello delle zone induttrici. Nelle zone effettrici, l'antigene può accedere per endocitosi attraverso le cellule epiteliali o attraverso le così dette "zone" o "unioni strette". La captazione e presentazione si realizza mediante macrofagi, cellule M o linfociti B ed i passi successivi seguono la procedura prima descritta. La capacità di induzione di una risposta immunitaria a livello delle mucose, richiede generalmente una maggior quantità di antigene ed a volte un maggior numero di immunizzazioni che non a livello sistemico, soprattutto nelle immunizzazioni per via orale. |
|||||
| Questo
è dovuto alle molteplici alterazioni e degradazioni enzimatiche che
subiscono gli antigeni attraverso questa via. Questo è un buon
meccanismo per la difesa dell'animale, ma è da tenere
in considerazione al momento di produrre vaccini orali. Anche
se, come vedremo nel capitolo 8, esistono diverse strategie per indurre
una buona risposta immunitaria anche per via orale. Tuttavia, è
generalmente più facile, indurre immunità nelle mucose respiratorie
mediante una immunizzazione orale, che
indurre la risposta immunitaria nelle mucose intestinali mediante
immunizzazione nasale. |
||||||
|
|
L'IgA
(capitolo 4)
svolge un ruolo molto importante nella risposta
immunitaria delle mucose. La sua configurazione a forma
di dimero o tetramero le permette di disporre da 4 a 8 siti di unione
con l'antigene. Questa caratteristica la rende estremamente
efficace contro i diversi antigeni batterici, mediante reazioni
del tipo ADCC, dato
che l'IgA non è battericida. L'IgA presenta grande capacità
di neutralizzare alcuni virus, inclusi all'interno delle cellule
epiteliali. E' l'unica immunoglobulina capace di agire all'interno
di una cellula, ma soprattutto la sua
principale attività nella difesa delle mucose
è quella di evitare l'aderenza di batteri e virus sulla superficie
dell'epitelio. L'IgA, può quindi agire in tre luoghi
e forme diverse. Da una parte, può unirsi all'antigene nel lume intestinale per
evitare l'aderenza del virus e/o batteri sulla superficie dell'epitelio;
può agire all'interno degli enterociti, e
per ultimo, nel liquido tissutale. |
|||||
|
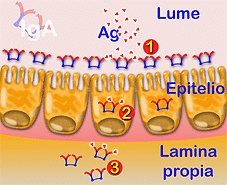
SCHEMA DI AZIONE DELL' IgA. L' IgA può agire a livello del lume intestinale, evitando l'aderenza sull'epitelio di virus e/o batteri (1), all'interno degli enterociti neutralizzando virus (2), e per ultimo, nel liquido tissutale (3). |
||||||
|
Un'ulteriore prova dell'importanza che questa immunoglobulina presenta nei meccanismi di difesa della specie suina, è rappresentata dal fatto che nella scrofa l'85% delle cellule che contengono immunoglobuline nella lamina propria dell'intestino presentano IgA. |
||||||
|
||||||